The Linear Oasis: Ecohydrology of Red Shank
Skip Navigation Links
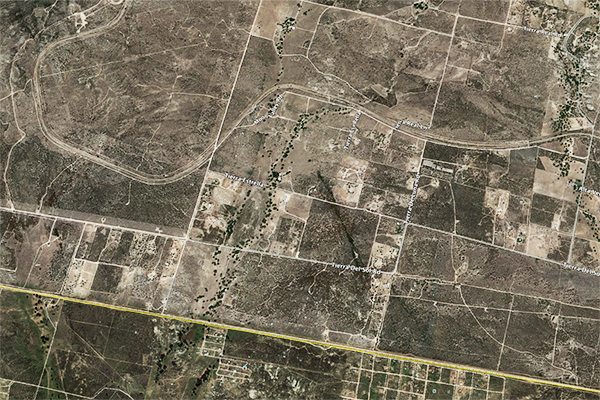
1. A LINEAR OASIS The dictionary has several definitions of oasis. One: A fertile spot in the desert, where water is found. Two: A small area in a desert that has a supply of water and is able to support vegetation. Three: An area where groundwater lies close enough to the surface to form a spring. These definitions point to the four features defining an oasis: A relatively confined spot, an arid climate, the presence of water, and the presence of vegetation. The community of Tierra del Sol, in southeast San Diego County, in California, has an arid climate, with mean annual precipitation averaging 15 in. There is, however, high variability in annual rainfall and associated environmental moisture. The record shows that annual rainfall has been as low as 4 inches in 2001-02 and as high as 31 inches in 1992-93 (Ponce et al., 2000; Ponce, 2006). Significantly, the region is underlain by a shallow fractured rock aquifer featuring an extensive system of fractures and dikes. The prevailing ecosystem is the chaparral, which is present in Southern California, extending into Baja California, Mexico. The region contains the ideal conditions for an oasis: A limited area, a desert climate, a shallow aquifer, and upland vegetation. Figure 1 shows a color-enhanced aerial view of a section of Tierra del Sol, near the Morning Star Ranch, a 358-acre rural property owned by Ed and Donna Tisdale. Vegetation is shown in bright red color. Two distinct features are noted:
Figure 2 shows a recent (2015) satellite image showcasing the linear forest. It is clearly seen that this forest would not be there if it were not for the presence of sufficient moisture to sustain it. In all probability, the linear forest overlies an unusually large fracture through which water and moisture are convected from the underlying aquifer to the surface. The objective of this study is to substantiate the relation between aquifer fractures and linear vegetative features, with particular focus on the Tierra del Sol watershed.
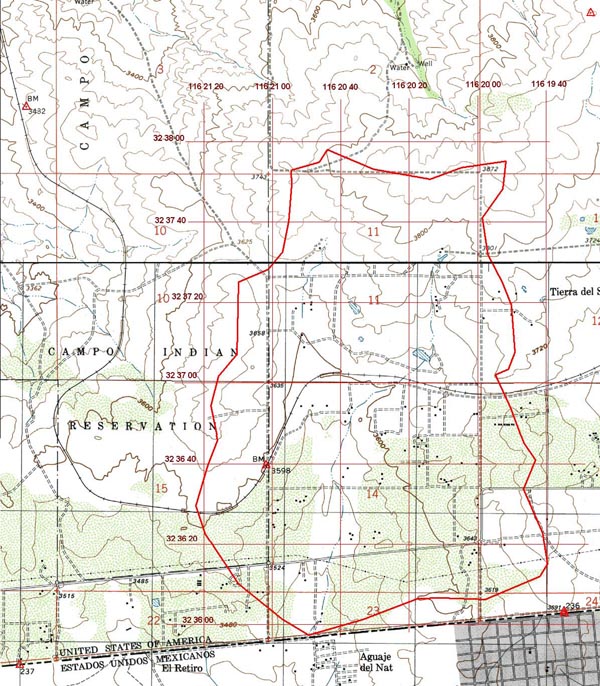
2. THE TIERRA DEL SOL WATERSHED The Tierra del Sol watershed comprises 1,760 acres near the southeast corner of San Diego County (Fig. 3). It drains gentle slopes and foothills of the California Peninsular Range in a predominantly southern direction, with numerous rock outcrops, dikes, and springs. The land use is a mixture of the native chaparral ecosystem, albeit with a substantial presence of phreatophytes along well defined moisture paths and, notably, a few mesophytes, such as the blue elderberry (Sambucus mexicana).
The anthropogenic land uses include
ranch clearings, rural housing, and other dwellings. The main watercourse is an ephemeral stream
which flows predominantly in a southern direction, crossing into Mexico at 32° 35" 56.89' N and
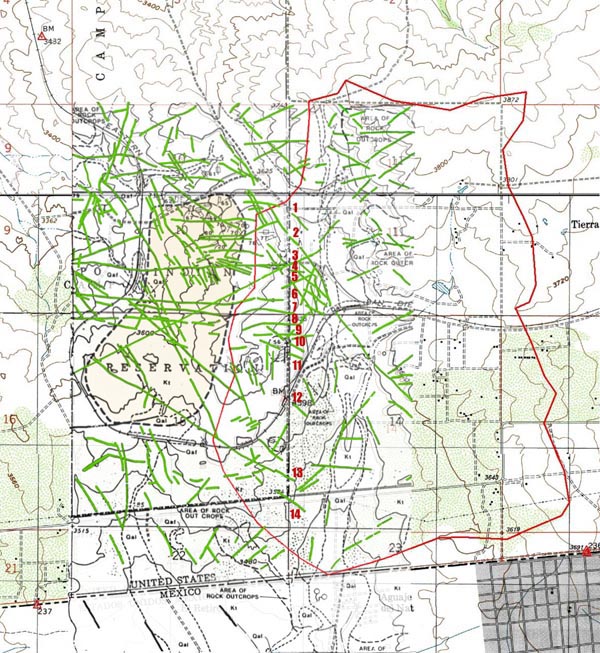
Existing studies have revealed the extensive fracture system in the underlying rock aquifer. Figure 4, prepared for the proposed Campo landfill project (1991), documents the presence of many photogeologic lineaments along the watershed's western boundary (Ponce, 2006). The lineaments, highlighted in green, depict changes in type and density of vegetation, reflecting the presence of fractures and other discontinuities in the underlying rock. Unlike the flow of water through an alluvial aquifer, where diffusion is the predominant process, transport through a fractured rock aquifer is dominated by advection.
A striking feature of the Tierra del Sol watershed, and one possibly related to the fractures, is the striking number of white, mostly pegmatitic, seemingly vertical dikes that cut through the underlying aquifer. These dikes are formed by quartz and feldspar-rich magmas left over in the final stages of cristallization of the granitic rocks (Ponce, 2006). One significantly major dike (Fig. 5) has been documented to be 1,465-ft long and about 6-ft wide (Fig. 6).
The Tierra del Sol watershed has its highest point on its northern upland ridges, at elevation 3,880 ft. Its lowest point is at its mouth on the U.S.-Mexico border, at elevation 3,440 ft. Thus, the total watershed relief is 440 ft. The numerous springs located in upland areas are a good indication of the hydraulic connection between groundwater and surface water.
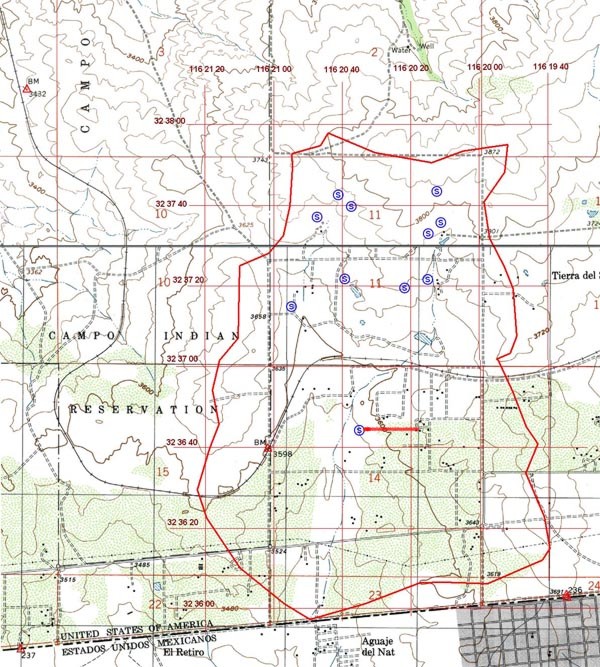
Figure 7 shows the location of eleven (11) springs
which drain mostly headwater slopes at elevations above 3,700 ft.
The southernmost spring shown in Fig. 7
is located precisely at the western extremity of the large dike (portrayed with a red line) shown in
3. RED SHANK vs CHAMISE Two closely related chaparral species, chamise (Adenostoma fasciculatum) and red shank (Adenostoma sparsifolium) are significantly represented in the Tierra del Sol watershed. Notably, these two shrubs are the only two species in their genus. Chamise is widely distributed throughout California and Baja California, with a range of nearly 800 miles. In contrast, red shank occurs only in four isolated places in Central California, Southern California, and Baja California as co-dominant with chamise, with a range of only 300 miles (Marion, 1943; Sampson, 1944; Hanes, 1965). Although chamise and red shank have overlapping geographical distributions, Beatty (1984) has noted that they do not grow in close association within a given stand. Red shank is commonly found on all aspects of gentle slopes and foothills of inner mountain ranges. Pure stands of red shank can be found in only 3% of the area in which red shank is the dominant species (Marion, 1943).
Chamise and red shank, although congeners, are not at all alike in appearance. Stands of chamise are dull, of dark-green color and uniform in appearance. The mature chamise plant is a medium-sized shrub 2 to 8 ft tall, with sparse leaf litter. In contrast, stands of red shank are distinctive, spreading above the general level of the chaparral. Red shank is a tall, round-topped arborescent plant 6 to 20 ft tall, with thick naked reddish stems and considerable leaf litter, 0.5 to 2 inches in depth (Hanes, 1965). The difference in amount of leaf litter between chamise and red shank may be attributed to their different water affinities (moisture availability and, consequently, actual evapotranspiration).
There is a substantial difference in the rooting habits of the two Adenostoma species. Chamise, even though less than half the average size of red shank shrubs, has a more agressive root system, possessing stronger roots with greater length and vertical penetration, appearing to be well adapted to moisture deficiency. On the other hand, the roots of red shank are massive, smaller, weaker, and are more likely to spread laterally rather than vertically. Chamise blooms and sets seed best, with low seed viability, following winters of ample rainfall. In contrast, red shank blooms well, with low seed set and viability, regardless of the amount of rainfall the previous winter (Hanes, 1965). Red shank appears to dominate over chamise on sites with higher moisture content and nutrient availability (Beatty, 1984). Hanes (1965) noted that although both Adenostoma species resumed growth in early winter, chamise displayed a sudden flush of growth in the spring, whereas red shank grew more steadily, with continued growth throughout the summer and fall. Chamise flowered in April and May, whereas red shank flowered abundantly in August. While chamise has an apparent enforced dormancy in the fall, red shank has been known to experience substantial autumnal growth, suggesting a sustained moisture source. Red shank appears to violate several definitions of sclerophyllous plants. First, it remains physiologically active during summer drought; thus, it is drought tolerant without being drought dormant (Hanes, 1965). Secondly, its shallow root system suggests that its moisture for summer growth must come from the top layers of the substrate. Therefore, red shank seems to be a type of shrub well adapted to drought conditions, but lacking the obvious morphological characteristics suggesting such adaptability (Shreve, 1934). The water affinities of red shank appear to lie in between those of the xerophytes, which are well adapted to drought, and those of the mesophytes, which habitually require a more sustained moisture source (Ponce, 2006). Thus, the link between red shank and groundwater exfiltration in an arid environment may be envisioned.
4. STUDY METHODOLOGY Soil moisture measurements were performed in two sets of paired locations: (a) the target point, located close to the center of the range delimited by the vegetative lineament, and (b) the control point, located outside of this range. To be meaningful, the measurements are performed periodically, at regular intervals, for a period of two years. The objective is to show that there is a moisture gradient between the target point and the control point, wherein the soil moisture in the target point is significantly greater than that of the control point. This explains the presence of the vegetative lineament, closely tied to a sustained moisture source originating in groundwater exfiltration through the linear rock fractures. The following field strategy was adopted:
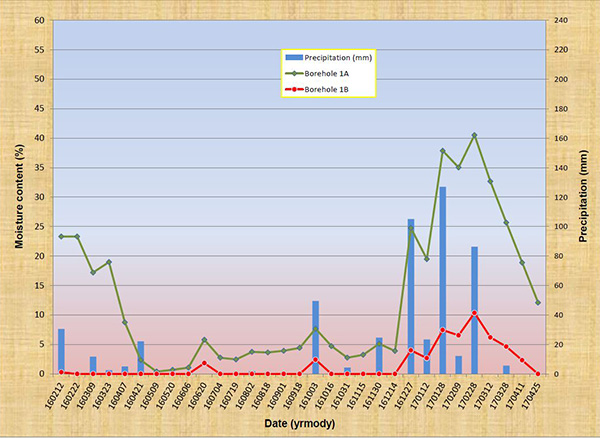
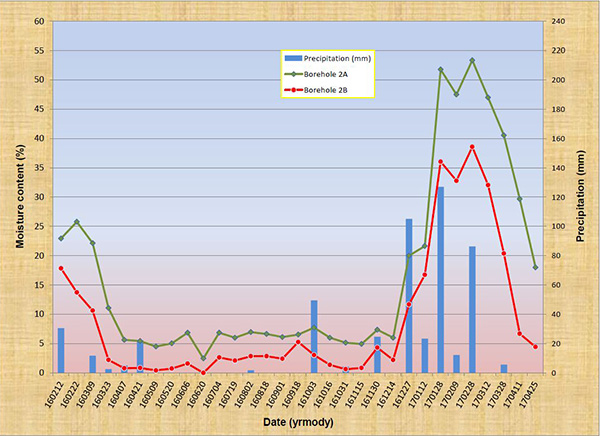
5. ANALYSIS Figure 15 shows a plot of the results, covering the 15-month period from 160212 to 170425. Figure 15 (a) shows the depth-averaged moisturecontent (%) in Boreholes 1A and 1B, while Fig. 15 (b) shows the depth-averaged moisture content (%) in Boreholes 2A and 2B. The A boreholes (target) are close to the fractured-rock lineament, while the B boreholes (control) are not. Intervening precipitation (mm) is shown to facilitate analysis. The plots clearly show that the moisture content in the target points (A) (green lines) is substantially higher than that of the control points (B) (red lines). At no time was the soil moisture content in B greater than its corresponding value in A. These results confirm that there are substantial and measurable differential moisture gradients in Tierra del Sol, attributable to exfiltration from the underlying fractured rock aquifer. To complement the analysis, temporal jumps in soil moisture content, in response to significant rainfall events, are documented.
6. CONCLUSIONS The present study warrants the following conclusions:
ACKNOWLEDGEMENTS
The financial support of Ed and Donna Tisdale, Backcountry Against Dumps,
and the people of Boulevard, in southeast San Diego County,
California, is gratefully acknowledged.
REFERENCES
Beatty, S. W. (1984). Vegetation and soil patterns in Southern California chaparral communities. In B. Dell, editor, Medecos IV:
Proceedings, 4th International Conference on Mediterranean Ecosystems, August 13-17, 1984, The Botany Department, University of Western Australia, Nedlands, Australia, 4-5.
Marion, L. H. (1943). The distribution of Adenostoma sparsifolium. American Midland Naturalist, 29(1), January, 106-116.
Hanes, T. L. (1965). Ecological studies of two closely related chaparral shrubs in Southern California. Ecological Monographs, 35(2), 213-235.
Ponce, V. M., R. P. Pandey, and S. Ercan. 2000. Characterization of drought across climatic spectrum.
ASCE Journal of Hydrologic Engineering, Vol. 5, No. 2, April, 222-224.
Ponce, V. M. 2006. Impact of the proposed Campo landfill on the hydrology of the Tierra del Sol watershed.
Online report, May.
Ponce, V. M. 2014. Effect of groundwater pumping on the health of arid vegetative ecosystems.
Online report, December.
Sampson, A. W. (1944). Plant succession on burned chaparral lands in Northern California. University of California Agricultural Experiment Station Bulletin 685, Berkeley, California, March, 144 p.
Shreve, F. (1934). The problems of the desert. The Scientific Monthly, 38(3), March, 199-209.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||














| 170627 15:00 |
| PDF file: download Adobe Acrobat Reader |